

Валерий Сойфер. Арифметика наследственности
Глава 11. Ступенчатый аллеломорфизм
Горячие точки гена
За несколько лет до опытов со скьютами американец Альтенбург доказал, что разные гены мутируют по-разному: одни быстрее, другие медленнее. Когда были найдены различные «центры» одного гена, появилась возможность определить, как часто мутируют отдельные участки гена: 11 мутаций гена скьют — это солидный материал для измерений. Да к тому же в поле зрения исследователей был еще и соседний, ген ахета.
Чтобы доказать неодинаковую частоту мутаций в разных участках — центрах гена скьют — Дубинин сделал такой расчет. Он подсчитал, сколь часто отдельные центры изменились во всех изученных мутациях, а затем построил график. По оси абсцисс были нанесены по порядку все центры гена скьют, а по ординате откладывалась суммарная частота мутирования каждого центра.
И вот на график ложатся точки. В центре скьюта они вырисовывают крутую дугу вверх: здесь участки, наиболее подверженные изменениям. У краев гена наклоны дуги спадают вниз: тут скорость изменений падает. Та же закономерность была отмечена и в гене ахета.
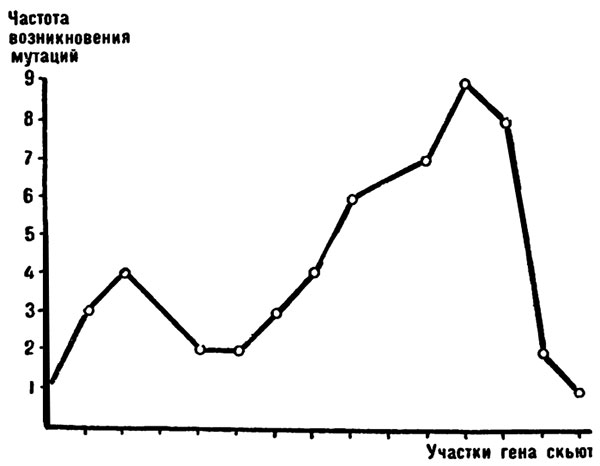
В различных участках гена скьют была обнаружена разная частота мутирования.
Есть участки, где мутации почти не возникают, зато в других местах гена мутаций бывает в пять—десять раз больше.
Спустя четверть века, уже в конце пятидесятых годов физик Сеймур Вензер, с головой ушедший в биологические опыты, изучал бактериофаги. Видимо ничего не зная об экспериментах Дубинина, он повторно обнаружил ту же закономерность. Отдельные точки гена и в его опытах изменялись чрезвычайно интенсивно,— Вензер назвал их «горячими точками».
Открытие Дубинина сразу же попало в центр внимания биологов. В истории генетики оно закрыло страницу, на которой ген описывался как исключительно устойчивая и достаточно простая единица наследственности: «Ген — единица функции, кроссинговера и мутации». Эта удобная и ставшая привычной для большинства генетиков формула потерпела первое крушение. Теперь уже нельзя было сказать, что ген мутирует как целое. Ступенчатый аллеломорфизм развеял это заблуждение.
Конечно, такой оборот дела не мог удовлетворить всех. Первым против теории гена Дубинина выступил признанный всем миром авторитет — генетик Рихард Гольдшмидт. Его устные высказывания чередовались с письменными, но смысл их оставался одним: они отрицали новую теорию гена. Как известно, одними восклицаниями в научной полемике многого не достигнешь. Чтобы критика стала действенной, надо что-то противопоставить теоретическим взглядам своего противника, объяснить его же факты, но с совершенно других позиций. В 1931 году появилась статья Гольдшмидта, в которой он выставил свою физиологическую, теорию наследственности. Сделана она была, надо сказать, на живую нитку. Чтобы хоть как-то обосновать свою точку зрения, Гольдшмидту пришлось прибегнуть к массе допущений и предположений.
Другим критиком теории дробимости гена выступил Генрих Фризен. Он изучал мутации гена уайт (белые глаза) у дрозофил и получил серию мутантов, сходных со ступенчатыми мутациями скьют-ахета. Данные Фризена вполне соответствовали теории Дубинина, хотя сам автор этого не заметил и решил обрушиться на теорию дробимости гена.
Однако спустя короткий промежуток времени он был вынужден признать сплошным недоразумением свое утверждение о неделимости гена. Согласившись с основными моментами теории ступенчатого аллеломорфизма, Фризен вместе с Дубининым выступил со статьей «Невозможность объяснения случая ступенчатых аллеломорфов скьют с точки зрения физиологической теории наследственности Р. Гольдшмидта». Впоследствии Гольдшмидт согласился с теорией дробимости гена.
Новая теория завоевывала себе всё больше сторонников. В 1959 году генетики М. Демерец и Ф. Хартман опубликовали большой обзор по генетике микроорганизмов. Они сочли необходимым начать его с опытов, проведенных совсем не на микробах, а на дрозофиле. Подробно описав эксперименты со скьютами, Демерец и Хартман подчеркнули, что эти опыты предвосхитили последующие достижения генетики. Стали той платформой, от которой отталкивались позднее ученые.









