

Валерий Сойфер. Арифметика наследственности
Глава 10. Ген в классическом понимании
Эффект положения
Еще в 1925 году Стёртевант, изучая мутации Bar у дрозофилы, отметил, что при перенесении гена из одной хромосомы в другую его активность меняется. Явление он назвал эффектом положения. С тех пор генетики не раз замечали, что проявление гена зависит от того, в соседство с какими генами он попадет. Но каждое такое наблюдение не могло дать окончательного ответа на вопрос о том, одним ли эффектом положения вызывается изменение работы гена.
В 1933—1935 годах Н. П. Дубинин и Б. Н. Сидоров в своих исследованиях обнаружили пример эффекта положения.
Они изучали ген жилкования крыльев, локализованный в маленькой, четвертой, хромосоме дрозофилы. Ген оказался рецессивным, и когда обе гомологичных хромосомы содержали его, одна из жилок крыла мух прерывалась.
Как ни мала четвертая хромосома, но кроссинговер в ней может произойти, и однажды в результате перекреста в четвертую хромосому по соседству с этим геном внедрился небольшой участок третьей хромосомы. Клетки тела содержат по паре каждой хромосомы, и парная четвертая хромосома мухи несла нормальный доминантный ген. По всем законам генетики, муха должна была иметь крылья со всеми жилками.
Однако как только по соседству с рецессивным геном поселился кусочек из другой хромосомы, произошло невероятное: рецессивный ген начал проявляться даже при наличии доминантного партнера. На него доминантный ген почти не действовал — жилка прерывалась.
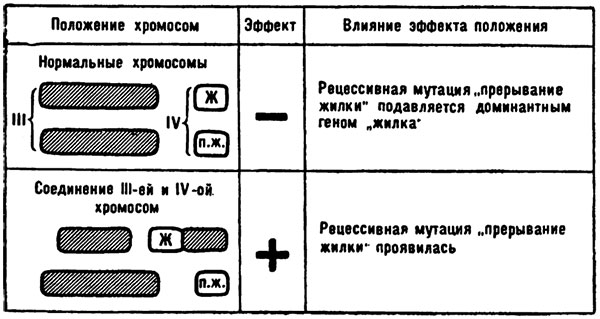
Эффект положения, описанный Н. П. Дубининым и Б. Н. Сидоровым в отношении гена «прерывание жилки».
Тонкий генетический анализ показал, что никакого изменения в самом рецессивном гене не произошло. Переход из подавленного состояния в активное связан был только с изменением окружения гена. Объяснить это явление иначе как эффектом положения Дубинин и Сидоров не могли. Правда, чтобы окончательно увериться в реальности этого эффекта, следовало осуществить еще один опыт: обменять лишний кусочек, оказавшийся в четвертой хромосоме, то есть восстановить прежнюю структуру. Если при этом рецессивность гена восстановится, тогда всякие сомнения относительно виновности эффекта положения отпадут.
С этой задачей ученые отлично справились. В третьей хромосоме есть ген хэйри. Он вызывает у мух образование дополнительных щетинок, но лишь в том случае, когда присутствует в обеих хромосомах. Другими словами, это рецессивный ген. Но стоило третьей хромосоме с геном хэйри присоединить кусочек четвертой хромосомы, как этот ген активно заработал. «Забыв» о том, что он рецессивный, ген хэйри начал диктовать мухам свои условия, и на теле их образовались дополнительные щетинки. Напоминаю, что его партнером в другой, третьей хромосоме так и остался ген, прежде подавляющий проявление хэйри.
Смена соседей привела к смене рецессивности на доминантность.
С помощью кроссинговера ученые поменяли местами доминантный хайри и бывший до этого рецессивным ген. Как только хэйри из третьей хромосомы с добавком четвертой хромосомы попал в нормальную третью хромосому, он мгновенно «утихомирился». Доминантный ген стал, как и прежде, подавлять проявление вернувшегося на место кочующего гена, а тот ничем не проявлял своего переменчивого нрава. Явление изменения активности гена и зависимости от перемены места в хромосомах могло считаться доказанным.
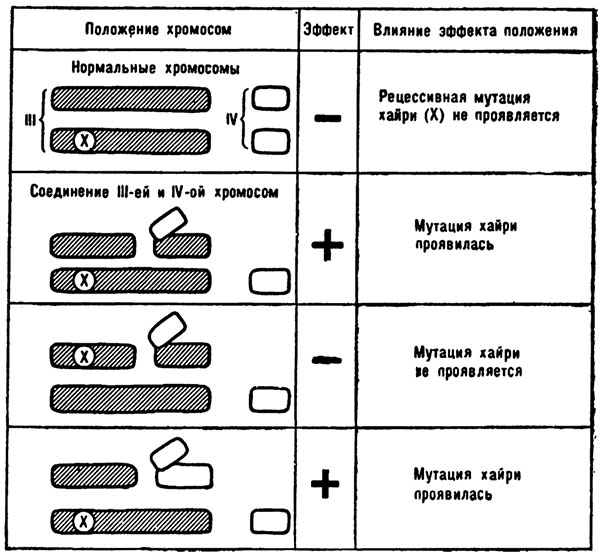
Эффект положения гена хайри.
Итак, к началу тридцатых годов были сформулированы основные положения теории гена. Согласно данным многочисленных экспериментов, ученые установили:
- каждый ген является элементарной структурной единицей наследственности;
- ген занимает вполне определенное место на хромосоме;
- ген определяет развитие вполне определенного признака (или группы признаков);
- может находиться в одном из аллельных состояний и мутирует как целое;
- при кроссинговере передается как единое целое, никогда не дробясь.
Многим тогда казалось, что эти положения незыблемы. Но схема эта не выдержала проверки времени. В 1929 году теории неделимости гена был нанесен непоправимый удар.









