

Валерий Сойфер. Арифметика наследственности
Глава 16. Генетический код
Сеймур Бензер и его r-II мутанты
Современные ученые думают, что область
изучения бактериального вируса представляет
собой великолепную площадку для игр,
на которой могут порезвиться серьезные дети,
любящие задавать глубокомысленные вопросы.
Макс Дельбрюк
Сеймур Вензер был физиком-теоретиком, но интересовался и другими науками. Какое-то седьмое чувство подсказало ему, что центральное место в науке постепенно завоевывает биология. Тогда он решил бросить все свои теоретические изыскания в физике и переучиться на биолога. Но биология велика. Опять-таки каким-то чутьем он понял, что следует заняться микробиологией. Однако, в отличие от многих физиков, полагавших, что стоит только приложить свои знания «хитрых физических материй» к примитивным биологическим понятиям, как «сезам откроется», Вензер почти десять лет потратил на то, чтобы овладеть специальностью микробиолога. И эти десять лет для него не прошли зря.
Он принялся за изучение бактериофага Т4, поражавшего бактерию — кишечную палочку. Вензер настолько хорошо узнал повадки этого фага, что мог рассказать о нем немало интересного. Кишечные палочки бывают разными. Имеются линия S, линия К, линия В и еще с десяток других линий, но для нас важны именно эти три. Обычный (дикий) тип фага Т4 мог прикрепляться к клеткам каждой из этих линий и впрыскивать туда свою ДНК. При росте на поверхности твердых питательных сред бактерии образовывали сплошную пленку, или бактериальный газон. Когда к бактериям добавляли фаг, он «выедал» на поверхности газона круглые мелкие пятна-плешинки, или, как их называют в бактериологии, бляшки. Время, уходившее на это, было устойчивым признаком фагов. Но иногда среди фагов наблюдались мутанты, названные от английского слова rapid (быстрый) — г-мутантами: r-мутанты образовывали бляшки быстрее, чем нормальные дикие фаги, и бляшки эти были крупнее и четче.
Бензер и занялся изучением r-мутантов фага Т4. Прежде всего ему удалось разбить их на три типа, которые он обозначил римскими цифрами I, II и III. Основанием для такого разделения служило то обстоятельство, что мутанты разных типов могли поражать лишь некоторые из линий кишечной палочки. Так мутанты r-II вообще не давали на газоне К никаких бляшек, а на газоне В образовывали большие бляшки.
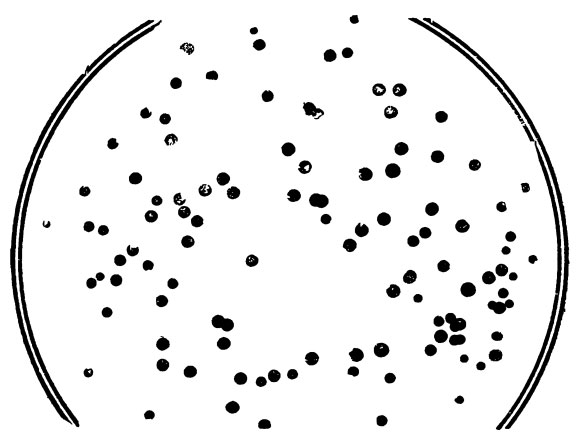
Бляшки бактериофагов
Этими r-II-мутантами Бензер и воспользовался. Привлекала простота их отбора. Допустим, на чашках с газоном кишечной палочки линии В оказалось сто крупных г-бляшек. Чтобы выбрать из них r-II-мутанты, достаточно посеять фаги из каждой бляшки на чашки с газоном кишечной палочки К, и все фаги, которые не дадут бляшек, будут r-II-мутантами. Благодаря этому Бензер быстро отобрал несколько сот r-II-мутантов фага Т4.
Первое, что он решил проверить: а нельзя ли получить рекомбинантов при скрещивании двух r-II-мутантов? Любая рекомбинация тут же дала бы о себе знать, ибо при рекомбинации участок с мутацией I перешел бы из хромосомы А в хромосому В, и получилась бы одна частица фага с двумя мутациями и одна нормальная частица. Мутантная частица не росла бы на линии К, зато нормальная частица тут же дала бы бляшку.
Согласно классической теории гена, рекомбинации в одном гене происходить не могли. Правда, Дубинин и Соколов, Льюис и Гриин смогли осуществить кроссинговер внутри гена. Но было неясно, приложимы ли их выводы ко всем генам. Однако первые же эксперименты Бензера привели его к успеху. Рекомбинанты образовывались. Снова фаги показали себя как нельзя лучше. Помните, в опытах Льюиса, для того чтобы получить кроссинговер в одном гене, понадобилось провести 100 000 скрещиваний, затратить огромный труд. А с фагами все было просто. 100 000 частиц можно было смешать с бактериями в одной пробирке, а затем высеять всю смесь на одну чашку Петри. Минимум труда — максимум успеха. Бензеру ничего не стоило получить массу рекомбинантов.
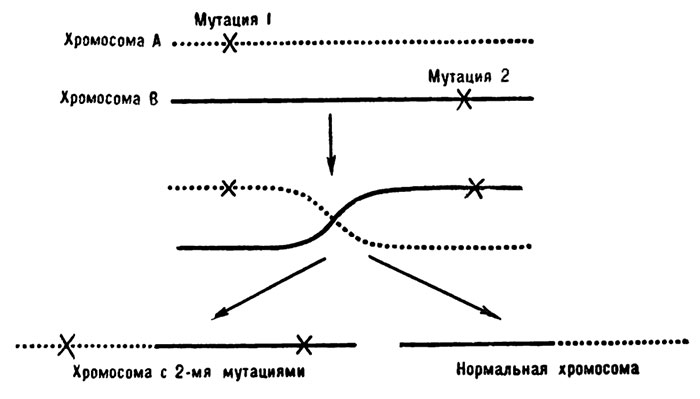
Схема рекомбинации
А потянув за эту ниточку, он смог распутать и весь клубок. Если внутри гена, управляющего проявлением одного признака, образуется не одна (как считали раньше), а много мутаций и если эти мутации могут рекомбинировать, то, установив частоту осуществления рекомбинаций, можно определить взаимное расположение мутаций внутри гена, то есть картировать их. И Бензер, применив традиционный генетический метод «трех точек», сумел расположить изученные им несколько сотен мутаций r-II друг за другом на генетической карте. Правда, не обошлось без хитростей.
Если бы лет 20 назад генетику сказали, что кто-то за год картировал 400 мутантов, он ответил бы: «Это немыслимое дело. Вы ошиблись раз в 10—20». Действительно, процесс картирования, даже при всех положительных качествах фагов, сложен и трудоемок. Обычно полученный мутант можно вернуть к прежнему состоянию, если вызвать обратную мутацию в том же участке данного гена. Но часть мутантов, полученных Бензером, никогда не возвращалась к дикому типу. Внимательное изучение таких мутантов навело ученого на мысль, что они возникли в результате выпадения значительных кусков молекулы ДНК, а не замены одного основания другим. Такие мутации, как вы помните, называют делециями (нехватками). Они-то и позволили ускорить картирование других мутаций. Выбрав несколько таких делеций, Бензер прежде всего точно определил длину выпавшего участка ДНК (то есть длину делеций), а затем уже, пользуясь такими делециями как линейками известной длины, быстро измерил расстояние между всеми мутациями.
Когда-то высшим достижением генетики считалось создание карты генов. Теперь на повестку дня встала более тонкая проблема — составление внутригенной карты. Еще один шаг в глубь гена был сделан!









