

Валерий Сойфер. Арифметика наследственности
Глава 13. Знакомьтесь: ДНК!
Молекулярный механизм мутаций
В школе мы частенько занимались тем, что играли в «слова». С общего согласия выбиралось какое-нибудь слово, не очень длинное, и за пять—десять минут надо было путем перестановки букв этого слова сконструировать другие слова — чем больше, тем лучше. Нередко после перестановки единственной буквы с одного места на другое или выбрасывания всего одной буквы смысл слова менялся.
Эта игра может просто объяснить, как изменится генетическая запись в ДНК (то есть возникнет мутация) при переменах в чередовании оснований вдоль молекулы ДНК. Что произойдет, если из длинной смысловой последовательности оснований выбросить одну или несколько пар? Смысл исказится. То же самое будет, если в последовательность оснований вставить лишнюю пару.
Вставка или выкидывание одиночных «букв» приведет к возникновению так называемых точечных мутаций. Но выпасть может и значительный кусок ДНК, и чем он будет больше, тем существеннее может оказаться повреждение — мутация.
А всегда ли для возникновения мутаций требуется выпадение одной или нескольких пар оснований? Нельзя ли обойтись заменой всего лишь одного основания?
Из рассуждений Крика и Уотсона выходило, что если один из членов пары оставался на своем месте, то в пару к нему мог встать только его партнер. В противном случае из-за различной длины оснований связь между сахаром и фосфатами в сахаро-фосфатной цепи возникнуть не могла.
Другое дело, если бы удалось как-нибудь укоротить длинные основания или удлинить короткие. Авторам пришлось признать, что принципиально такая возможность имеется.
Вернемся к картинкам. Нарисуем двойную цепь ДНК с пустым местом против аденина.
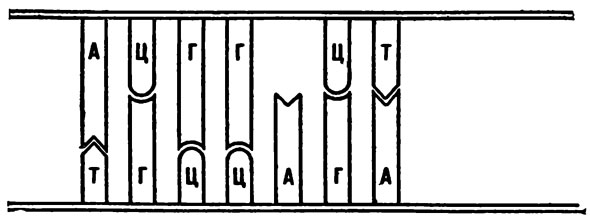
В результате нарушения структуры ДНК одно из оснований (на рисунке тимин) может выпасть из структуры нуклеиновой кислоты.
Как правило, на это место должен встать только короткий тимин. Но допустим, что каким-то образом на место тимина удастся втиснуться цитозину. До той поры, пока ДНК не начала делиться, ничего не будет заметно. Но после того как одиночные нити разойдутся и к ним начнут достраиваться парные нити, ситуация резко изменится. Та нить, в которой осталось нормальное основание (аденин), достроит себе нормальную нить, и двунитчатая ДНК будет абсолютно идентична родительской. А вот та нить, в которой оказался втиснувшийся чужак, поведет себя иначе. Ведь цитозин, по правилам, должен соединяться в пару с гуанином. Когда цитозин влезал не на свое место, ему приходилось свернуться. Но как только нити разошлись и ему стало вольготно, он «распрямился», и теперь уже в пару ему требовался гуанин. Однако если гуанин объединился с цитозином, в нормальном расположении оснований произойдет ошибка. Была пара А—Г, стала пара Г—Ц. Вот вам и мутация.
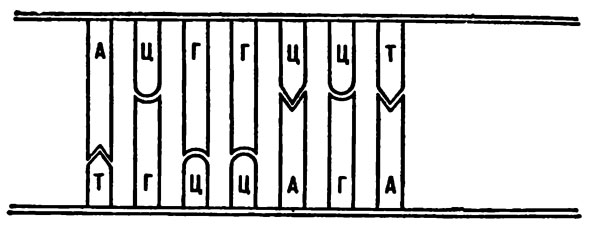
На выпавшее место в редких случаях может встать не тимин, как это полагается, а цитозин. Тогда возникнет мутация.
Мы начали с предположения, «что каким-то образом на место тимина удалось втиснуться цитозину». Возможно ли это? Вероятность подобной замены казалась ничтожно мала, до тех пор пока в 1956 году Донохю и Стент не предсказали один из путей, ведущих к ее осуществлению.
Чтобы познакомиться с принципом их рассуждений, нарисуем структурную формулу цитозина:
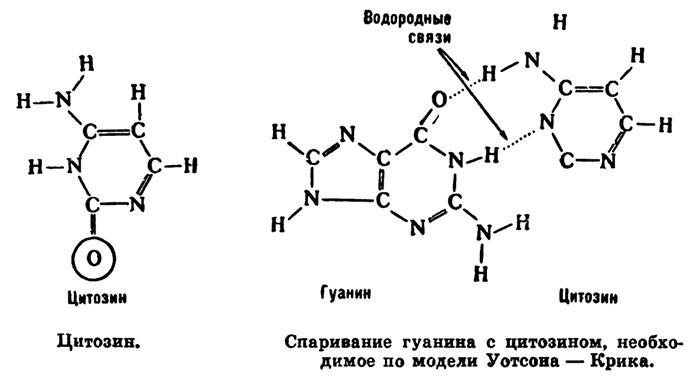
Спаривание гуанина с цитозином, необходимое по модели Уотсона — Крика.
Важно обратить внимание на атом кислорода, обведенный кружком. Если кислород находится в кето-форме, как изображено на рисунке, то цитозин соединится с гуанином.
Но кислород может быть присоединен к углероду и иначе, в энольной форме. Тогда не исключено, что у такого цитозина появится возможность соединиться с аденином: то есть как раз то, что и приведет к мутации.
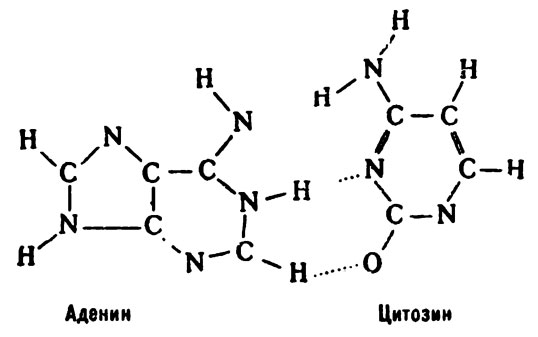
В результате ошибки спаривания цитозин может соединиться не с гуанином, а с аденином.
Эти предположения сбылись. В 1959 году американский исследователь Эрнст Фриз доказал, что такие неверные замены имеют место и приводят к мутациям.
Сразу же после открытия Крика и Уотсона в лабораториях всех стран мира начались работы по изучению причин, которые приводят к выпадению оснований или вставке лишних оснований в молекулу ДНК.
Число агентов, изменяющих структуру ДНК, быстро росло. Прежде всего удалось выяснить, что традиционные мутагенные факторы — рентгеновские лучи, ультрафиолетовые лучи, многие химические соединения — воздействуют вовсе не на белки, как считали генетики раньше, а в основном на ДНК. Начало развиваться новое направление науки — изучение факторов, влияющих на нуклеиновые кислоты. Удалось найти много веществ, которые были отдаленно похожи на аденин, гуанин, цитозин или тимин и потому могли замещать их в цепи ДНК во время ее удвоения, что приводило к изменению правильного чтения ДНК на неправильное. Список таких «троянских коней» растет с каждым днем.
Заканчивая этот раздел, можно с удовлетворением отметить, что гипотеза Крика и Уотсона столь же великолепно справилась и с объяснением молекулярных причин мутаций. Предложенная ими структура ДНК позволяла осуществляться мутациям. Но, как всегда, решив одну задачу, ученые столкнулись со множеством других. Перед ними встала новая, гораздо более трудная проблема. Надлежало объяснить, как ДНК управляет жизнью клетки, как ее приказы передаются из ядра в цитоплазму, кто выполняет роль курьеров. Но... этого пока Крик и Уотсон объяснить не могли. «В настоящее время нам неясно, каким образом она осуществляет эту функцию»,— писали они. Понадобилось еще несколько лет, прежде чем и эта загадка была решена.
Не могли Крик и Уотсон представить доказательства и в пользу своего основного постулата — о механизме продольного разделения нитей ДНК и последующей их достройки. Верность этого предположения установили спустя пять лет.
В 1958 году Месельсон и Сталь поставили следующий эксперимент. Вырастили клетки бактерий на среде, где все атомы азота были представлены своим тяжелым изотопом N15. В ДНК клеток, выросших на такой среде, все атомы азота стали тяжелыми, и ученые могли теперь легко отличить такую тяжелую молекулу ДНК от ее легких собратьев в других бактериях. Это очень пригодилось Месельсону и Сталю.
Клетки с тяжелыми молекулами ДНК перенесли в среду с легким изотопом азота N14. Наступила пора поделиться клеткам. Прежде всего начали расходиться нити ДНК. Теперь каждая нить должна была пристроить себе вторую половину. Но в качестве строительного материала клетки могли предложить только легкие блоки. Вот тут-то и настал главный этап опыта. Если модель Крика и Уотсона верна и нити ДНК не рассыпаются, не разваливаются, то одна половина вновь образующейся двойной полимерной молекулы останется тяжелой, а вторая половина будет достроена из легких атомов азота. Именно это и зарегистрировали приборы.
Ученые продолжили наблюдение. В следующем акте деления неравноценные половинки ДНК должны были снова удвоиться, после чего половина двунитевых молекул ДНК стала бы целиком содержать легкие атомы, а половина по-прежнему осталась бы гибридной. Это предсказание также полностью сбылось, подтвердив экспериментально верность гипотезы Крика и Уотсона о строении ДНК и ее генетической роли. После опытов Месельсона и Сталя уже уверенно можно было говорить о теории Крика и Уотсона.









