

Валерий Сойфер. Арифметика наследственности
Глава 14. Бактерии - папы, бактерии - мамы
Полярность переноса хромосом
В первой своей работе Хейс обнаружил явление, которое он назвал полярностью переноса генов. Хейс работал с бактерией кишечной палочки. И он не замечал, чтобы все известные ему гены передавались женской клетке. Всегда одни гены передавались чаще, другие реже, но были и такие, которые крайне редко оказывались в женских клетках. Хейс предположил, что это следствие различного положения генов в хромосоме. Самый крайний ген передается первым, и потому его передача отмечается чаще. Гены, расположенные дальше от конца, попадут в женскую клетку позже. Из-за того что две такие клетки, сцепленные тонким мостиком, легко отрываются друг от друга, удаленные гены будут часто оставаться в мужской клетке. Но это предположение Хейса требовало выполнения одного жесткого условия: хромосома мужской клетки должна передаваться всегда одним концом. С чем связана такая неравноправность концов хромосомы, Хейс не знал.
А все было очень просто. Жакоб и Вольман определили, что хромосома начинает входить в женскую бактерию концом, противоположным тому, к которому присоединился F-фактор. В опыте Хейса все бактерии оказались потомками одной исходной бактерии. Естественно, что у этой линии порядок передачи генов был постоянным. В линии*, выделенной Кавалли-Сфорца, порядок передачи был также постоянным, но первым, вторым и т. д. передавались совсем другие гены. Это явление Хейс и назвал полярностью.
Доказательство полярности хромосомы, предложенное Вольманом и Жакобом, настолько красиво, что я не могу не рассказать о нем подробнее. Есть такой прибор смеситель Уоринга. Устройство его несложно. От электромотора идет ось, на которой укреплены лопасти. Что-то вроде машинки для приготовления молочного коктейля. Смесь с бактериями можно влить в стакан, а затем опустить в него стержень с лопастями. Нажим кнопки — и мотор пущен: в секунду-две все клетки, соединенные мостиками, будут разбиты. Этот смеситель Уоринга и использовали Вольман и Жакоб. Они соединили суспензии клеток Hfr и женских F-. Дали им возможность коныогировать и через минуту разрушили мостики между клетками. После этого изучили, какие гены успели перейти от клетки Hfr в клетку F-. Затем то же самое проделали через минуту, две и так далее, вплоть до двух часов.
Результат оказался блестящим. Тиминовый ген передавался на восьмой минуте, ген устойчивости к азиду натрия на восьмой с половиной минуте, лейциновый ген на девятой минуте, и т. д. По тому, какой ген успевал войти в клетку, можно было проверять часы. Вольман и Жакоб сделали рисунок, который обошел, вероятно, все биологические аудитории мира. Сбоку одной бактерии (около нее значок женской клетки  ) лежит другая бактерия (знак
) лежит другая бактерия (знак  показывает, что она мужская). Чуть повыше часы, минутная стрелка которых стоит на нуле. Рядом второй рисунок. Стрелка часов сдвинулась на минуту, а мужская клетка уже вплотную прикасается к женской. На следующем рисунке стрелка ушла на пять минут, и между клетками образовался проход, в который вдвинулся конец хромосомы мужской клетки, на котором отмечено 0, то есть начало хромосомы, и чуть отступя А (ген А). Итак, рисунок за рисунком дают представление о том, как с течением времени всё новые и новые участки мужской хромосомы проникают внутрь женской клетки. Вот прошел час, и клетки оторвались друг от друга, и за это время половина мужской хромосомы вошла в F- клетку.
показывает, что она мужская). Чуть повыше часы, минутная стрелка которых стоит на нуле. Рядом второй рисунок. Стрелка часов сдвинулась на минуту, а мужская клетка уже вплотную прикасается к женской. На следующем рисунке стрелка ушла на пять минут, и между клетками образовался проход, в который вдвинулся конец хромосомы мужской клетки, на котором отмечено 0, то есть начало хромосомы, и чуть отступя А (ген А). Итак, рисунок за рисунком дают представление о том, как с течением времени всё новые и новые участки мужской хромосомы проникают внутрь женской клетки. Вот прошел час, и клетки оторвались друг от друга, и за это время половина мужской хромосомы вошла в F- клетку.
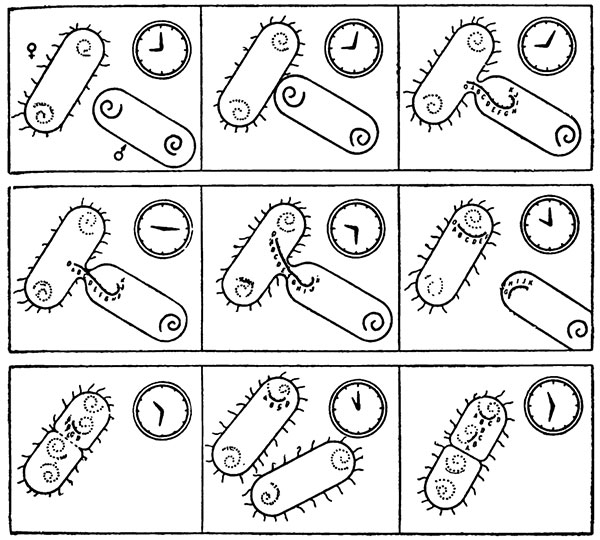
Рисунок Жакоба и Вольмана. Вхождение последовательных участков хромосом мужских клеток в женские можно отметить по часам.
Теперь окончательно выявилось коренное различие между высшими организмами и бактериями. Еще механизм отличия по полу как-то отдаленно совпадал. У высших организмов причина была в лишней хромосоме (X) или в разных хромосомах (X и У), у бактерий также можно было за лишнюю хромосому принять F-фактор. Но на этом сходство кончалось. При оплодотворении у высших ядра сперматозоидов сливались с ядрами яйцеклеток, а хромосомы объединялись в одной клетке, формируя зиготу. У бактерий клетки не сливались. Ядра не объединялись. Просто возникал мостик между клетками, и в эти ворота осторожно вплывала хромосома, на другом конце которой прикрепился маленький фактор пола. Передача хромосомы почти никогда не осуществлялась целиком. Что успело, то и втиснулось, а затем мостик рвался, разрывая и мужскую хромосому. Мужская клетка с обрывком своей хромосомы отходила в сторону.
Исследования Вольмана и Жакоба прочно утвердили тезис Хейса о полярности переноса и доказали, что только Hfr клетки могут передавать свои признаки. Клетки со свободными факторами плодовитости не могут этого сделать. Но теперь для решения загадки, почему хромосома в клетках со свободными F-факторами не передается, оставалось сделать всего один шаг.









